

{kind=link}
All Alteromonas strains help long-term survival of Prochlorococcus below N hunger
Earlier analysis confirmed that Prochlorococcus, and to some extent Synechococcus rely upon co-occurring heterotrophic micro organism to outlive varied varieties of stress, together with nitrogen hunger [33, 34, 42, 43]. On the first encounter between beforehand axenic Prochlorococcus and Alteromonas (E1), all co-cultures and axenic controls grew exponentially (Fig. 1B, C). Nonetheless, all axenic cultures confirmed a speedy and principally monotonic lower in fluorescence beginning shortly after the cultures stopped rising, reaching ranges under the restrict of detection after ~20–30 days. Not one of the axenic Prochlorococcus cultures have been capable of re-grow when transferred into recent media after 60 days (Fig. 1C). In distinction, the decline of co-cultures quickly slowed, and in some instances was interrupted by an prolonged “plateau” or second development stage (Fig. 1B). Throughout a number of experiments, 92% of the co-cultures contained residing Prochlorococcus cells for at the very least 140 days, which means that they could possibly be revived by switch into recent media. Thus, the flexibility of Alteromonas to help long-term N hunger in Prochlorococcus was conserved in all analyzed strains.
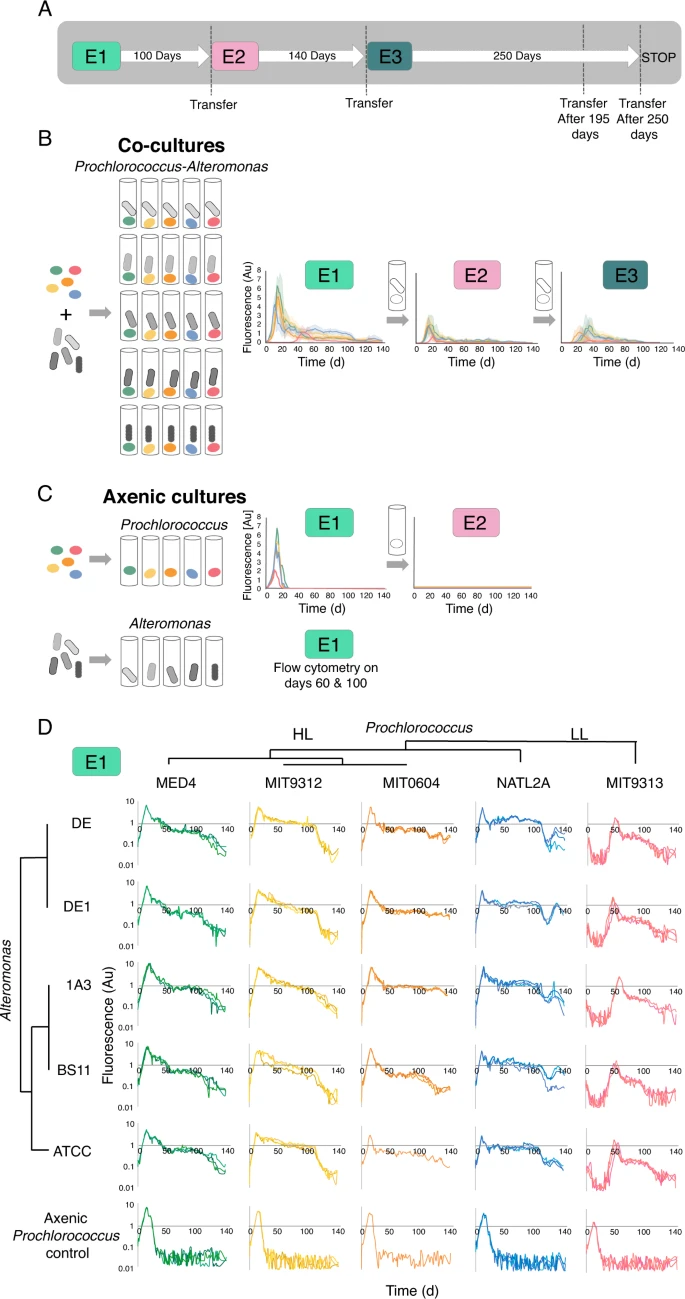
A Schematic illustration of the experimental design. One ml from Experiment E1 was transferred into 20 ml recent media after 100 days, beginning experiment E2. Experiment E2 was equally transferred into recent media after 140 days, beginning experiment E3. Extra experiments replicating these transfers are described in Supplementary Fig. S1. B Overview of the expansion curves of the 25 Prochlorococcus-Alteromonas co-cultures over three transfers spanning ~1.2 years (E1, E2 and E3). Outcomes present imply + normal error from organic triplicates, coloured by Prochlorococcus pressure as in panel D. C Axenic Prochlorococcus grew exponentially in E1 however did not develop when transferred into recent media after 60, 100, or 140 days. Axenic Alteromonas cultures have been counted after 60 and 100 days, as their development can’t be monitored sensitively and non-invasively utilizing fluorescence (optical density is low at these cell numbers). D Excessive reproducibility and strain-specific dynamics of the preliminary contact between Prochlorococcus and Alteromonas strains (E1). Three organic replicates for every mono-culture and co-culture are proven. Notice that the Y axis is linear in panels B, C and logarithmic in panel D. Au: arbitrary models.
It has beforehand been proven that Prochlorococcus MIT9313 is initially inhibited by co-culture with Alteromonas HOT1A3, whereas Prochlorococcus MED4 is just not [12, 32]. This “delayed development” phenotype was noticed right here too, was particular to MIT9313 (not noticed in different Prochlorococcus strains) and occurred with all Alteromonas strains examined (Fig. 1D). MIT9313 belongs to the low-light tailored clade IV (LLIV), that are comparatively distant from different Prochlorococcus strains and differ from them in a number of physiological points together with the construction of their cell wall [44], using totally different (and nitrogen-containing) appropriate solutes [45], and the manufacturing of a number of peptide secondary metabolites (lanthipeptides, [46, 47]). LLIV cells even have bigger genomes, and are predicted to take up a better variety of natural compounds similar to sugars and amino acids [48]. It’s intriguing that particularly this pressure, which has larger predicted metabolic and regulatory flexibilities [49], is the one one initially inhibited in co-culture with Alteromonas.
Variations in co-culture phenotype are associated to Prochlorococcus and never Alteromonas strains and happen primarily through the decline stage
Whereas co-culture with all Alteromonas strains had a serious impact on Prochlorococcus viability after long-term hunger, there was no vital impact of co-culture on conventional metrics of development similar to maximal development price, maximal fluorescence, and lag part (apart from the beforehand described inhibition of MIT9313; Fig. 2A–C). Nonetheless, a visible inspection of the expansion curves prompt delicate but constant variations within the form of the expansion curve, and particularly the decline part, between the totally different Prochlorococcus strains within the co-cultures (Fig. 1D). To check this, we used the expansion curves as enter for a principal element evaluation (PCA), revealing that the expansion curves from every Prochlorococcus pressure clustered collectively, no matter which Alteromonas pressure they have been co-cultured with (Fig. 2D). The expansion curves of all high-light tailored strains (MED4, MIT9312, and MIT0604) have been comparatively comparable, the low-light I pressure NATL2A was considerably distinct, and the low-light IV pressure MIT9313 was a transparent outlier (Fig. 2D), in keeping with this pressure being the one one initially inhibited in all co-cultures. Random forest classification supported the commentary that the expansion curve shapes have been affected extra by the Prochlorococcus fairly than Alteromonas strains, and in addition confirmed the visible commentary that a lot of the options differentiating between Prochlorococcus strains occurred throughout tradition decline (random forest is a supervised machine studying algorithm defined in additional element in Supplementary Textual content S2; see additionally Supplementary Fig. S2). Thus, co-culture with Alteromonas impacts the decline stage of Prochlorococcus in co-culture in a means that differs between Prochlorococcus however not Alteromonas strains.
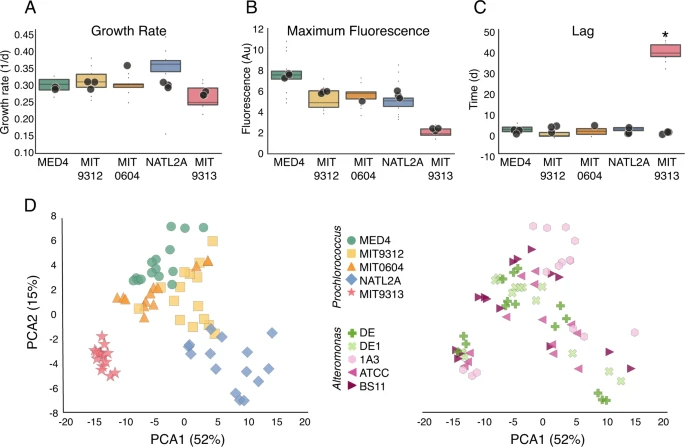
A Development price, B Most fluorescence, and C length of lag part throughout experiment E1. Field-plots characterize imply and seventy fifth percentile of co-cultures, circles characterize measurements of particular person cultures of the axenic controls. The one vital distinction between axenic and co-cultures is within the size of the lag part for MIT9313 (Bonferroni corrected ANOVA, p < 0.001). D PCA ordination of the expansion curves coloured by Prochlorococcus (left) and by Alteromonas (proper) strains. The expansion curves cluster by Prochlorococcus pressure (Adonis take a look at, F(4,68) = 42.3, R2 = 0.71, p = 0.001) and solely marginally by Alteromonas pressure (Adonis take a look at, F(4,68) = 2.29, R2 = 0.11, p = 0.017). Au arbitrary models.
We subsequent requested whether or not the phenotypes of interplay, which have been noticed when excessive cell densities of Prochlorococcus and Alteromonas interacted for the primary time (E1), have been maintained after the cells had lived collectively in co-culture for prolonged intervals. We subsequently continued to switch the co-cultures into recent media over a number of further transfers, carried out 40–200 days after the preliminary inoculations. In complete, fluorescence measurements can be found for a cumulative interval of 380 days, which the cells spent in co-culture (Fig. 1A; Supplementary Fig. S1). The flexibility of Prochlorococcus to outlive long-term N hunger, the clustering of the expansion curves by Prochlorococcus however not Alteromonas strains, and the outcomes of the random forest classification, have been all reproduced in subsequent transfers (Fig. 1B; Supplementary Figs. S2, S3; Supplementary Textual content S3). These observations are thus sturdy to the cumulative time the organisms have been interacting and the cell densities of each organisms when transferred to new media (see under).
Variations within the carrying capability counsel totally different modes of interplay
Whereas Alteromonas clearly help Prochlorococcus, by enabling it to outlive long-term N hunger, is the reciprocal interplay additionally synergistic? Do Prochlorococcus improve the expansion of Alteromonas, and does the interplay have an effect on the general carrying capability of the system, outlined right here as the flexibility to effectively make the most of the limiting useful resource (nitrogen)? To reply these questions, we used the circulation cytometry cell counts of Prochlorococcus and Alteromonas on days 60, 100, and 140 of experiment E1 to deduce the nitrogen (N) biomass of every inhabitants grown alone or in co-culture (Fig. 3; Supplementary Desk S1; see Strategies and Supplementary Textual content S4 for the calculations and caveats).

A Development curves of experiment E1, arrows displaying the times the place cell numbers have been counted by circulation cytometry (see additionally Supplementary Desk S1). Axenic curves proven are from all Prochlorococcus strains. Thick strains characterize imply fluorescence. B Whole calculated N biomass of the cultures in E1 on days 60 and 100 in μmol N/L. Dashed line signifies complete out there N in co-cultures. C Log2 Fold Change (log2FC) of Alteromonas N biomass relative to axenic Alteromonas controls on days 60 and 100. Asterisks point out statistically vital FC (Bonferroni corrected ANOVA p < 0.05). ALT Alteromonas, PRO Prochlorococcus, Au arbitrary models.
The general carrying capability of the co-cultures was larger than the axenic Alteromonas cultures, and far larger than the axenic Prochlorococcus (Fig. 3B). On day 60, the imply carrying capability of the co-cultures was 2–3 occasions larger than that of the Axenic Alteromonas (69 ± 35 in contrast with 32 ± 22 μmol N/L), suggesting that the heterotroph benefited from carbon fastened by the phototrophic Prochlorococcus accomplice. Certainly, most of this mobile N was discovered within the Alteromonas cells (76 ± 13%, see Supplementary Textual content S4 for a sensitivity evaluation). The flexibility of axenic Alteromonas to outlive within the absence of natural matter from Prochlorococcus is no surprise, as an Alteromonas pressure distantly associated to those studied right here, AltSIO, can make the most of a big fraction of the labile natural materials present in pure seawater used to make the expansion media [50]. In distinction, in axenic Prochlorococcus cultures solely a small fraction of N within the system was present in cell biomass (~0.01 μmol N/L). This probably displays the lack of all Prochlorococcus strains to recycle natural nitrogen misplaced because of exudation or cell lysis (Fig. 3B).
Mutual synergism was not noticed throughout all pressure combos. Whereas some Prochlorococcus strains (MED4, MIT0604, and NATL2A) supported considerably larger Alteromonas N biomass in comparison with the axenic management (log2FC 1.3 ± 0.6), co-cultures with MIT9312 and MIT9313 resulted in comparable or decrease Alteromonas biomass (log2FC −0.2 ± 0.9) (Fig. 3C). Due to this fact, on day 60, among the interactions have been mutually synergistic whereas in different instances Prochlorococcus don’t help Alteromonas and should even compete with it. The 2 “non-mutually-synergistic” Prochlorococcus strains belong to totally different ecotypes however have been remoted from the identical drop of water from the Gulf Stream [51]. In all co-cultures the Prochlorococcus inhabitants benefited from the presence of Alteromonas (log2FC 10 ± 4 in N biomass in comparison with axenic controls).
In distinction to day 60, after 100 days basically the entire interactions have been mutually synergistic, with Alteromonas supporting the expansion of all Prochlorococcus strains (log2FC 10 ± 1) and Prochlorococcus growing the Alteromonas biomass in all strains apart from BS11 (log2FC 3 ± 1.5) (Fig. 3C). This means that the mode of interplay (synergist verses competitors) might change temporally through the prolonged interval of N hunger.
On day 140 the carrying capability of the co-cultures declined additional (only one% of the N was in biomass). This means that the system is just not in regular state, with a sluggish but fixed discount in carrying capability. We speculate that that is pushed by the lack of bioavailable N from the system (i.e., a lot of the nitrogen is in a recalcitrant type that can’t be utilized by both accomplice). That is additional supported by the commentary that whereas the co-cultures after 140 days have been nonetheless alive and could possibly be transferred to new media, in a subsequent experiment, solely 16/30 cultures could possibly be transferred after 195 days, and solely 3/75 cultures could possibly be transferred after 250 days (Supplementary Fig. S5). MIT0604 (HLII) was the pressure almost definitely to outlive switch after these prolonged intervals and was additionally essentially the most ample Prochlorococcus pressure after 140 days (1.46 ± 1 μmol N/L MIT0604 biomass verses 0.26 ± 0.56 μmol N/L for all different strains). Whereas we don’t presently have an evidence for the upper survival of this pressure, it’s noteworthy that it’s the solely pressure to make the most of nitrate [52].
Modeling the impact of co-culture on Prochlorococcus mortality
On condition that the clearest impact of co-culture was on the decline part of the co-cultures, we requested whether or not we may quantify and mannequin the impact of Alteromonas on Prochlorococcus mortality. Whereas the expansion of micro organism has been extensively studied and modelled, the decline of bacterial cultures is way much less studied, and mortality is never represented in ecological or biogeochemical fashions of microbial dynamics [53]. Bacterial mortality has, nevertheless, usually been modelled within the context of meals security and genome evolution, utilizing both mechanistic or descriptive approaches [53,54,55,56,57]. We selected to concentrate on 4 of those beforehand described fashions that are comparatively easy and have a transparent organic interpretation (Desk 1). The exponential mannequin is the best and mostly used one, the place a continuing portion of the inhabitants dies over time [58]. The bi-exponential mannequin is barely extra complicated, representing two separate subpopulations locally, every with its personal demise price [55]. The Weibull mannequin is probabilistic, modeling a heterogeneous inhabitants with a various stress tolerance [53, 59], lastly, the harmonic mannequin employs a quadratic price of decline which is commonly related to predator-prey interactions or mobile encounter charges [58]. When becoming every of those fashions to the decline part of the expansion curves, the Weibull mannequin stands out because it has a low error for each axenic and co-cultures (Desk 1) in addition to in consequent transfers (Supplementary Desk S2), the bi-exponential mannequin is a greater match for the co-cultures however doesn’t characterize effectively the axenic ones. Primarily based on the Weibull mannequin, and assuming that tradition fluorescence is expounded to the variety of non-lysed cells within the media (Fig. S7), axenic Prochlorococcus cells die greater than ten-fold quicker than cells in co-culture (2-decimal discount time, td2, is 12.58 ± 3.85 days for axenic cultures and 316 ± 337 days for co-cultures). Comparable outcomes have been obtained with the bi-exponential mannequin (Supplementary Textual content S5).
Within the Weibull mannequin, the “form parameter” (n) represents the change over time (because the cultures decline) within the susceptibility of the bacterial neighborhood to emphasize. A form parameter above one represents an growing chance that cells will die as time will increase (e.g. as a result of accumulation of injury), whereas a form parameter under one means that, because the tradition declines, the cells change into extra resistant to break. Axenic cultures have excessive imply form worth of two.1 ± 0.9, suggesting an accumulation of cell harm resulting in growing demise price (Fig. 4A). In distinction, the imply form worth of co-cultures is considerably decrease and under 1 (0.4 ± 0.2, pupil t-test, p < 0.001), suggesting that in N hunger in co-culture the Prochlorococcus cells are acclimating over time to the nutrient stress circumstances.

A Weibull form (n) in Axenic Prochlorococcus (PRO) and in Co-cultures. In all axenic samples n > 1, in most co-cultures n < 1 (students t-test, p < 0.001). B Scatter plot displaying the reverse correlation between the overall N biomass of the co-cultures on day 60 and Weibull form (n). Pearson r = −0.33, p = 5e-3. Circles represent co-cultures with mutual synergistic interactions (i.e. Prochlorococcus strains MED4, MIT0604, and NATL2A), X characterize potential aggressive interactions (strains MIT9312, MIT9313). C Scatter plot displaying the correlation between the overall N biomass of the co-cultures on day 60 and Weibull root imply sq. error (RMSE). Pearson r = 0.64, p < 0.001. For the correlations with complete N biomass on day 100 see Supplementary Fig. S6. D–F Weibull mannequin match for chosen decline curves (FL). D Mutually synergistic co-culture of Prochlorococcus NATL2A and Alteromonas DE1. E Doubtlessly aggressive co-culture of Prochlorococcus MIT9312 and Alteromonas DE1. F Axenic Prochlorococcus NATL2A. FL fluorescence (arbitrary models). RMSE root imply sq. error.
Whereas the molecular and physiological mechanisms of Prochlorococcus adaptation are presently unclear, the Weibull form parameter decreases as the overall N in mobile biomass will increase, suggesting that the Prochlorococcus acclimation course of is expounded to the flexibility to recycle N between the precise Prochlorococcus and Alteromonas strains in co-culture (Fig. 4B). Thus, the speed of acclimation is larger within the co-cultures supporting excessive N biomass and mutually synergistic interplay (NATL2A, MED4, and MIT0604) in comparison with MIT9312 and MIT9313, the place Alteromonas don’t achieve from the interplay (0.36 ± 0.14 verses 0.52 ± 0.16, pupil t-test p < 0.001).
Whereas the Weibull mannequin is helpful in quantifying mortality charges and elevating the speculation that Prochlorococcus cells are acclimating to hunger over time, not one of the examined fashions was capable of absolutely seize the intricate dynamics of tradition decline (Fig. 4C, D). In most mutually synergistic co-cultures involving Prochlorococcus strains NATL2A, MED4, and MIT0604, tradition decline was not monotonic, and was interrupted by further development phases about 40–50 and 100 days after the cultures began declining (Fig. 4D). These latter development phases have been principally absent in co-cultures with MIT9312 and MIT9313 (RMSE 0.36 ± 0.1 verses 0.2 ± 0.1 within the different strains, pupil t-test p < 0.001). The correlation between N biomass and the secondary development phases (i.e., larger deviation from easy Weibull mannequin; Fig. 4C) counsel that these phases might also be associated to the flexibility of the interacting companions to recycle N via mutually useful metabolic interactions.
Conclusions and future prospects
Elucidating the mechanisms of microbial interactions requires well-characterized mannequin methods. Nonetheless, extending the insights from such fashions throughout the range of organisms and environmental circumstances stays difficult. Our outcomes from the extremely simplified system of a number of Prochlorococcus and Alteromonas strains present an necessary step in the direction of this purpose. Utilizing the wealthy data on interplay phenotypes current within the development and decline curves, we determine conserved and strain-specific aspects of those interactions. Regardless of the genetic variety throughout the Alteromonas strains studied [30], it was primarily the id of the Prochlorococcus pressure that decided the interplay phenotype. This manifests within the development and decline charges, within the form of the curve (primarily the decline part), within the quantity of N retained in biomass, and in whether or not the co-cultures are mutually synergistic or, probably, aggressive.
Beneath our laboratory circumstances, it’s probably that the mixed response of each interacting companions to nitrogen hunger underlies the dynamics of the long-term co-cultures, though different stressors similar to the rise in osmolarity/salinity or the buildup of waste merchandise can’t be dominated out [18, 34, 60]. This response is dynamic, as illustrated by the reproducible deviations of the fluorescence curves from the monotonic decline predicted by all fashions examined (“second development” levels; Fig. 4). Three totally different (non-mutually-exclusive) processes might underlie these dynamics. Firstly, it’s probably that one or each organisms modify their physiology or metabolism over time, for instance via the activation of stringent responses, utilization of N or C storage swimming pools, rewiring of metabolism to make the most of out there N sources, or activation of mechanisms similar to extracellular enzymes permitting the cells to entry beforehand unusable substrates (e.g. [61, 62]). Secondly, it’s attainable that there are “invisible” ecological dynamics underlying the noticed fluorescence curves, for instance cyclic modifications within the abundance of Alteromonas cells. Beneath such a state of affairs, speedy Prochlorococcus mortality may produce a rise in Alteromonas abundance, leading to degradation and remineralization of lifeless Prochlorococcus biomass and the discharge of assets that may drive subsequent Prochlorococcus development. Thirdly, each Prochlorococcus and Alteromonas populations could also be evolving, for instance via emergence of genetically distinct populations higher tailored to nutrient hunger (paying homage to the GASP phenotype described in E. coli and different micro organism [63]).
Why is it the id of the first producer (Prochlorococcus) fairly than the heterotrophic “recycler” (Alteromonas) that determines the end result of the co-culture? A-priori, it was cheap to imagine that the co-culture phenotype can be affected by the variations between the Alteromonas strains of their potential to degrade and make the most of polysaccharides and quite a lot of different natural molecules [30, 64]. We speculate that the elevated development of Alteromonas within the co-cultures in comparison with the axenic ones is fueled primarily by the provision of main biomass elements launched by Prochlorococcus as they die, similar to proteins, amino acids, and nucleotides. Such widespread macromolecules don’t require extremely specialised metabolic processes to degrade and make the most of, and therefore could be utilized by the entire Alteromonas strains [65]. It’s attainable that the variations between Alteromonas strains might manifest when extra complicated macromolecules can be found, e.g. from plant materials, or when the entire “simple to digest” (labile) natural matter has been utilized and solely complicated macromolecules stay [66]. These circumstances might not have been met in our experiments. It’s also attainable that co-culture with a extra various vary of heterotrophic micro organism, together with further Alteromonas species, would reveal extra pronounced variations within the impact of the heterotroph of the first producer. Equally, we presently have no idea why some Prochlorococcus strains help a mutually synergistic interplay with Alteromonas comparatively early through the long-term N hunger (day 60) whereas different strains don’t, and why at a later stage (day 100) virtually all interactions are mutually useful. We couldn’t determine any metabolic traits [11] clearly differentiating MIT9313 and MIT9312 (the “aggressive” strains) from the others, suggesting extra delicate variations exist between the Prochlorococcus strains within the natural matter they produce or of their response to N hunger (e.g. [67, 68]).
Our outcomes determine patterns within the interactions between clades of ample marine phototrophs and heterotrophs, below circumstances the place vitamins are scarce, and their availability probably depends upon recycling between phototrophs and heterotrophs. Whether or not or not such mechanisms could also be physiologically related within the oligotrophic ocean, a lot of which is N-stressed [40], stays to be examined. For instance, within the oceans, speedy turnover of Prochlorococcus cells because of grazing and viral lysis probably signifies that cells are, on common, youthful than these in laboratory cultures, which can have an effect on their mortality charges [69]. Moreover, stressors similar to phage an infection and grazing are lacking in laboratory cultures. It’s, nevertheless, noteworthy that the excessive heterotroph/phototroph biomass ratio noticed throughout long-term N hunger right here and in different research [18] is much like that of a lot of the open oligotrophic ocean (e.g. [70] and references therein). Moreover, Alteromonas might enable Prochlorococcus to adapt to gentle hunger [43] and to the presence of ROS (e.g. [71]), in addition to different stressors that may be encountered within the open ocean. The supportive position of Alteromonas can’t be taken as a right, because it additionally depends upon tradition circumstances, for instance CO2 concentrations [27].
The co-cultures didn’t attain a gentle state, and didn’t characterize a closed system. Thus, processes not represented in these simplified laboratory co-cultures, are essential to clarify the long-term stability over a long time of Prochlorococcus within the oceans [72]. Such processes may embrace multi-organism interactions, as pure communities are far more complicated than the laboratory co-cultures, in addition to oceanographic processes similar to nutrient injection via deep mixing. Extra typically, cell mortality is intimately linked with the quantity and sort of recycled natural matter, but the speed of mortality in pure communities is extremely unconstrained [73]. Therefore, higher illustration of mortality in mathematical fashions (e.g. using applicable mortality formulations) is probably going necessary for understanding biogeochemical cycles [73]. This will entail utilizing one of many “off the shelf” fashions introduced right here, with their limitations (e.g. the Weibull mannequin requires an estimate of the time of decline, whereas quadratic expressions for mortality are already utilized in such fashions [74,75,76]), or the event of recent fashions that higher hyperlink cell physiology, ecology, maybe genome construction, and mortality.